









20220909文献解读 | 动物发育不同阶段mRNA m5C修饰景观和新m5C修饰调控机制

研究背景和待解决的科学问题

RNA上目前发现存在着170多种不同类型的转录后修饰,所有已知的RNA种类都可以被修饰,其中rRNA和tRNA上的修饰最多。mRNA上的主要修饰类型包括m6A、m1A和m5C。m5C修饰是最早发现的RNA修饰之一,最初研究表明主要分布在tRNA和rRNA上,近期研究证实,真核生物的mRNA m5C修饰分布广泛,其可以维持mRNA的结构和稳定性、调控翻译过程等,但是在动物发育过程中m5C修饰动态变化、功能和进化仍然在很大程度上是未知的。
为了获得动物发育过程中mRNA m5C的景观、功能和进化关系,作者对跨越8亿年进化的6种动物物种进行bsRNA-seq【康测科技bsRNA-seq也可以做哦(~ ̄▽ ̄)~】,以构建不同发育阶段mRNA m5C修饰图谱,发现潜在的修饰调控机制。
研究内容
1. 母体mRNA m5C修饰水平在脊椎动物和无脊椎动物中都异常高
为了揭示脊椎动物和无脊椎动物物种发育过程中的m5C特征,作者对斑马鱼(Zebrafish)和黑腹果蝇(D.mel)样品进行了bsRNA-seq,涵盖所有发育阶段,如下图。
在斑马鱼和果蝇的早期胚胎阶段的3187和5021个mRNA中分别鉴定出6259和8974个外显子m5C修饰位点,这些mRNA主要从母体基因组转录出来。这些m5C位点数量在从母体到受精卵阶段后显著降低,而且在后续的发育阶段维持较低的水平,如下图。
随后作者关注其他动物特别是哺乳动物m5C修饰变化情况,并开展最能代表脊椎动物的分类:人、小鼠、非洲爪蟾和热带爪蟾的卵母细胞和早期胚胎细胞的bsRNA-seq。与斑马鱼和果蝇一样,结果显示这种母体→受精卵过程m5C修饰模式也保守地存在于两栖动物和哺乳动物中,如下图。
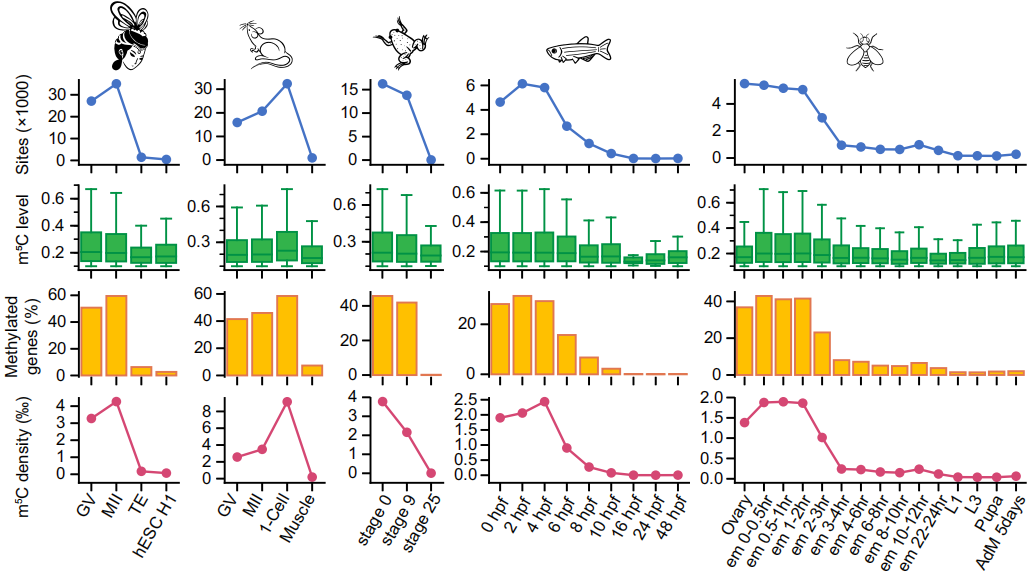
总体上,母体mRNA上m5C修饰密度比后续发育阶段mRNA要高出10-30倍,并发现在母体mRNA上存在大规模m5C修饰。
2. 在非人类物种中,母体mRNA m5C修饰主要由NSUN2添加
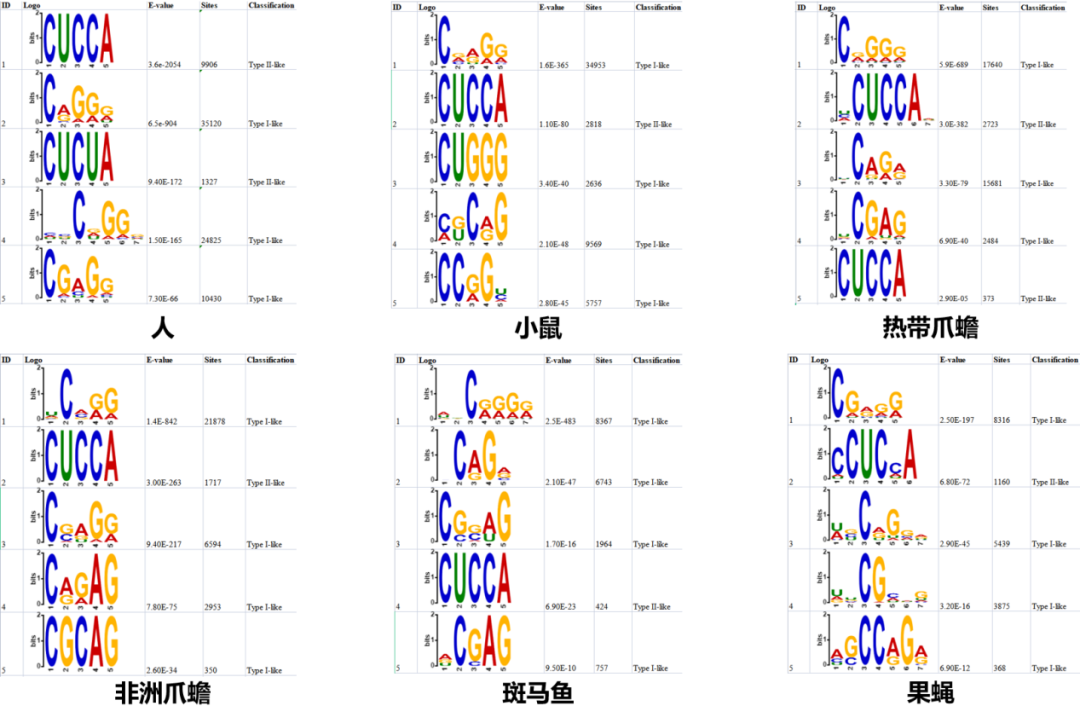
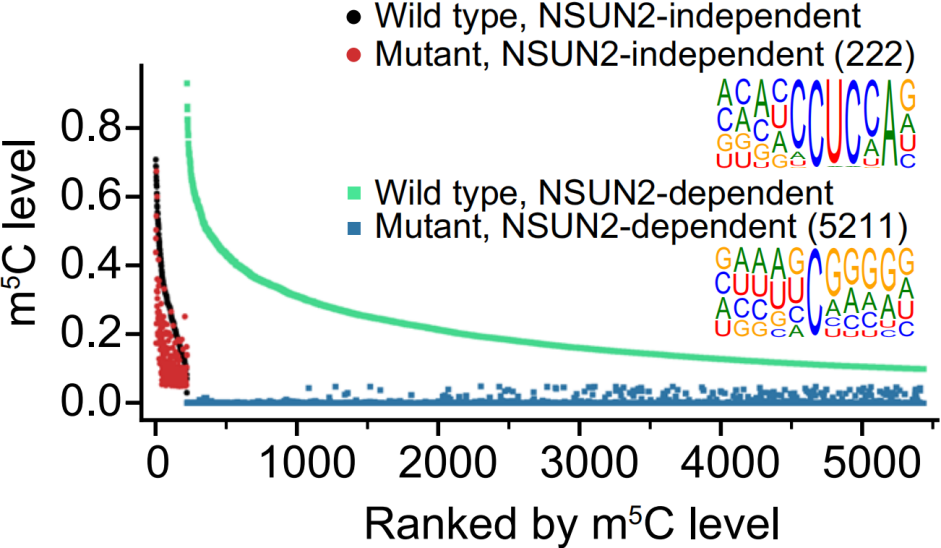
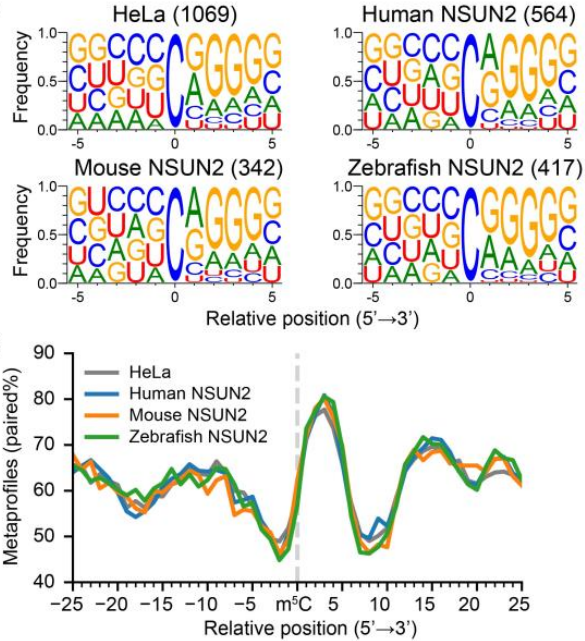
母体mRNA大规模m5C修饰是由哪种m5C Writer蛋白完成的,除了NSUN2是否还有其他Writer在起作用?为探究这两个问题的答案,作者对所有m5C修饰位点进行motif扫描,将motif归为两类,具体见下图。可以看到,CUCCA的motif在脊椎动物和无脊椎动物中都存在,比较保守。
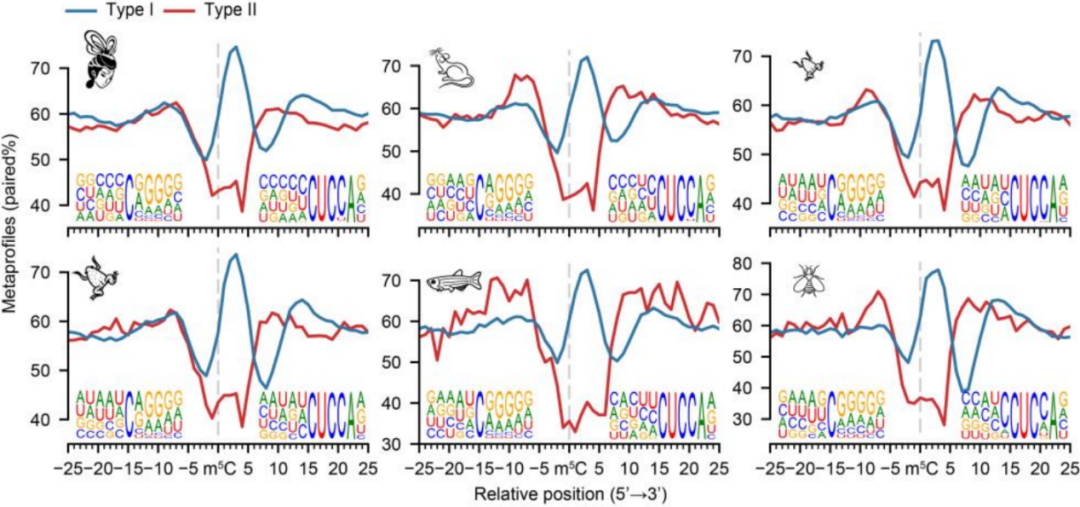
考虑到motif序列本身特征,作者根据其motif对母体mRNA m5C位点进行了分组,并检查了它们的结构特征,结果显示在所有物种中,具有两个motif的位点位于5'末端或发夹结构的环中,如下图。
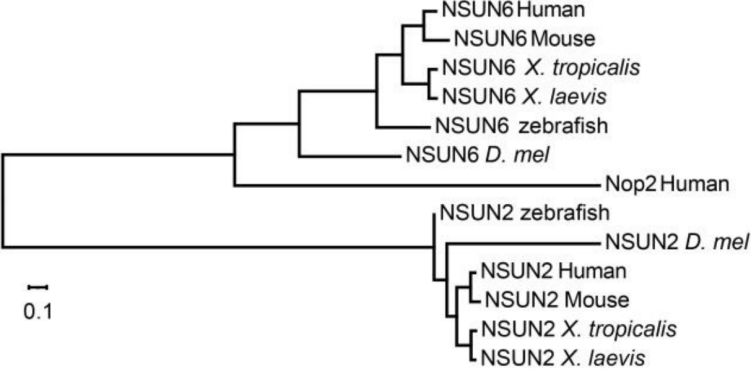
进化树分析显示所有物种NSUN2和NSUN6的保守性很高,如下图,说明主要是这两个Writer蛋白负责mRNA m5C甲基化。
随后,作者开始构建各物种母体mRNA m5C修饰景观:
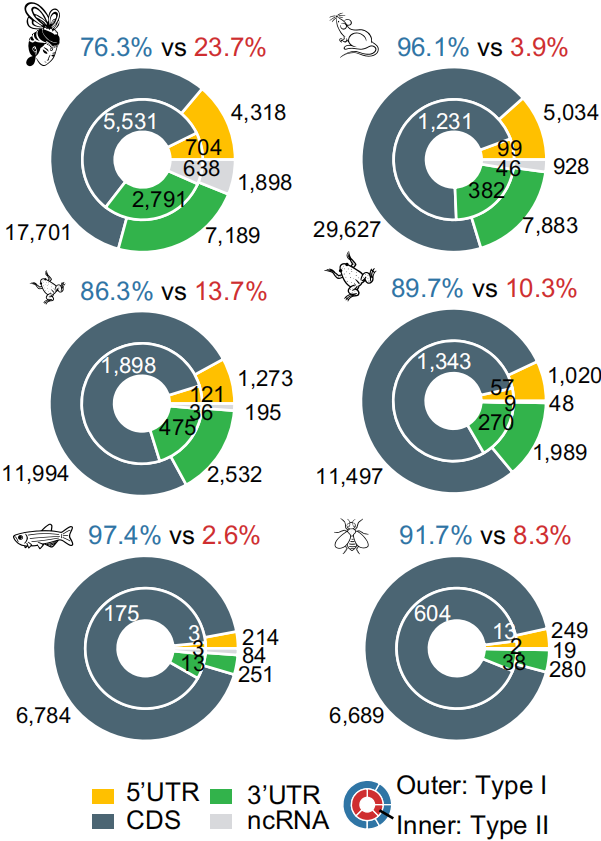
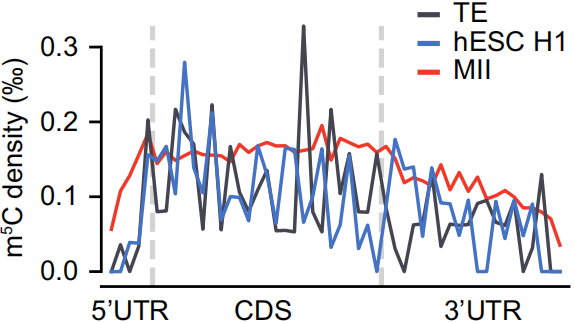
1. 果蝇和斑马鱼中~92% m5C修饰分布在CDS区;两栖和哺乳动物在双端UTR区域分别占比20.6%和36.8%,如下图;
2. 所有物种中Type I的motif占绝大部分(86%-97%),而人中Type II的motif有一定占比,如下图;
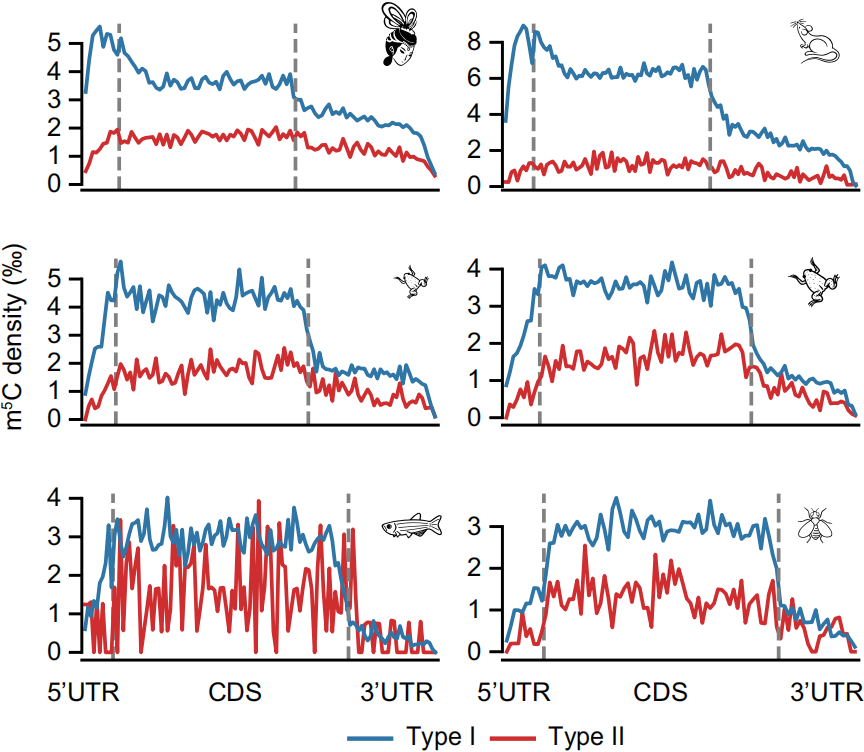
3. 与之前报道一致,Type I的m5C修饰在5’UTR末端富集,如下图;在所有物种中,Type I和Type II在CDS和3’UTR区分布模式相似,如下图。
为了阐明母体mRNA m5C甲基化的机制,作者分析公共数据库的RNA-seq数据,结果显示虽然NSUN2表达量非常高,但是也无法解释一些m5C修饰模式。母体mRNA m5C甲基化有两个特征:a. 修饰水平稳定;b. 大部分修饰导致翻译沉默并储存在细胞质中。因此作者假设NSUN2的细胞定位决定了其与mRNA相互作用的持续时间,可能与大规模m5C甲基化有关。
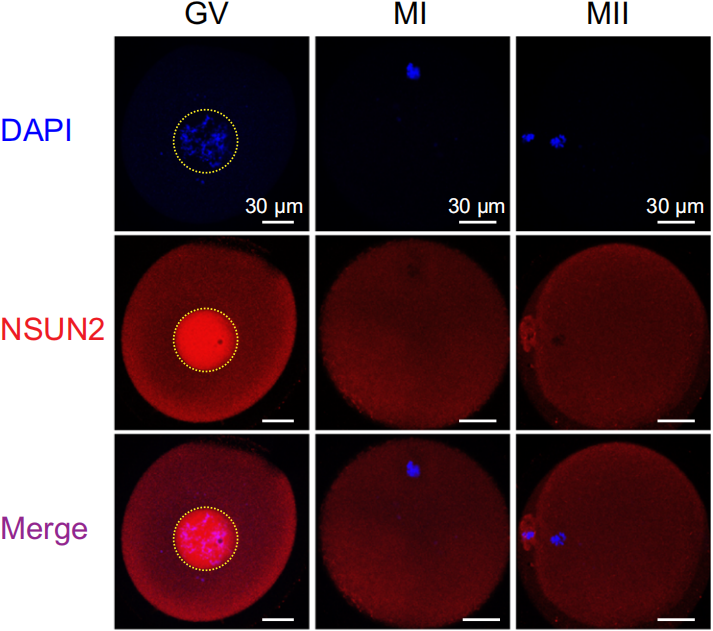
在果蝇卵母细胞中,NSUN2主要定位于分裂前期I(prophase I, PI)的细胞核中(如下图),随着分裂中期I(metaphase I, MI)核膜裂解,NSUN2释放到胞质中。
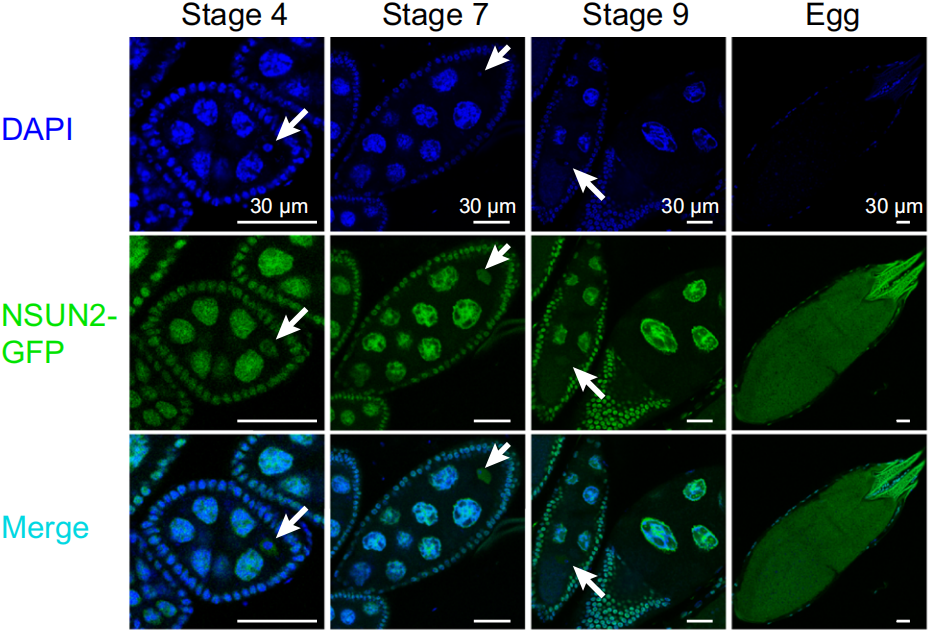
与果蝇不同,人和小鼠卵母细胞的NSUN2在PI期同时分布在细胞核和胞质中,如下图。与果蝇一样,核膜裂解后,NSUN2进入胞质。观察到这些结果后,作者提出细胞质中的NSUN2可以在卵母细胞成熟到受精过程中一直与mRNA相互作用,维持mRNA的m5C修饰水平。
3. Type I m5C位点的缺失会导致果蝇发育迟缓
作者观察到母体mRNA存在大规模m5C修饰,自然想到m5C可能调控母体mRNA的命运。作者首先想到缺失NSUN2后会造成什么影响,于是构建了NSUN2敲除的果蝇,结果显示NSUN2确实是主要负责母体mRNA m5C修饰(96%)的Writer。
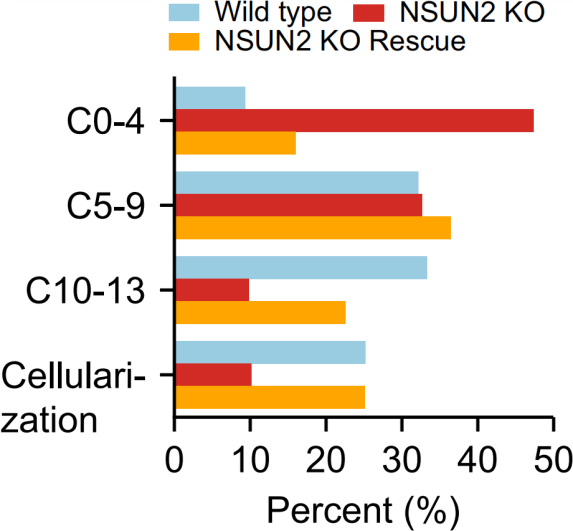
为了确定NSUN2在母体中的功能,作者将NSUN2 KO雌性果蝇与WT雄性果蝇进行杂交,大约35%的NSUN2 KO胚胎在向一龄幼虫发育时夭折。除此之外,WT胚胎通常从卵变成幼虫需要21.5 h,而NSUN2 KO卵平均延迟2 h。为了精确监测这一过程,作者统计处于胚胎发生不同阶段的卵的百分比。结果显示NSUN2 KO卵胚胎发生时间比WT卵更长,如下图。这种较慢的发育可以通过引入野生型,而不是催化域突变失活的NSUN2转基因而得到挽救,如下图。这些结果共同说明NSUN2是母体mRNA m5C修饰所必需的。
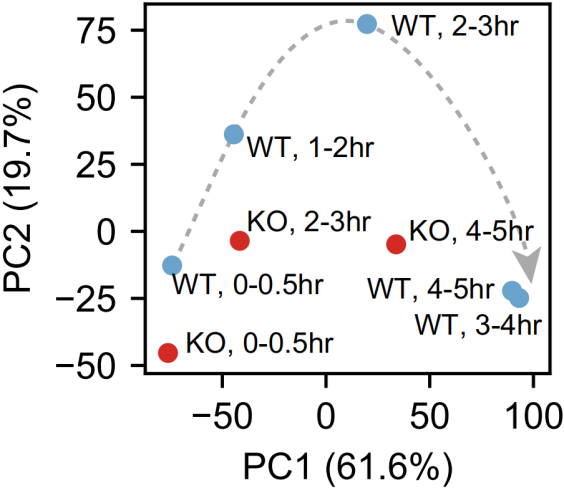
为了进一步验证在分子水平上观察到的发育迟缓,作者进行RNA-seq分析了不同时间点的WT和NSUN2 KO胚胎。PCA分析显示PC1是主要与胚胎发育相关的主成分,NSUN2 KO胚胎PC1值在2-3 h和4-5 h明显低于WT胚胎,如下图。
通过分析WT胚胎高表达基因,发现NSUN2 KO胚胎对应的这些基因明显延迟启动表达,如下图。这些结果共同说明NSUN2在母体mRNA m5C修饰方面的功能。
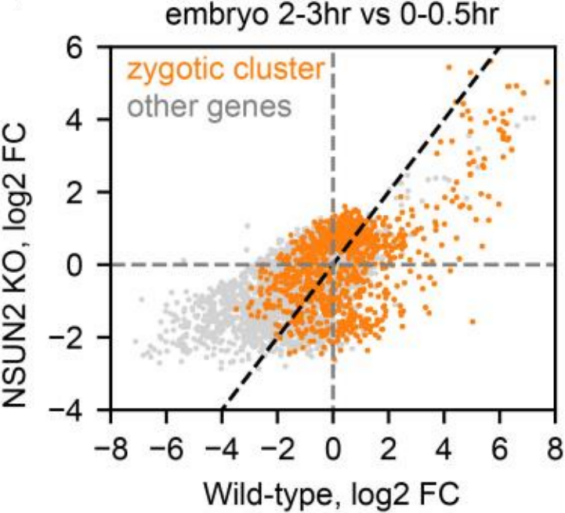
4. 新型顺式调控机制可以使高等动物5’UTR末端获得更多m5C修饰位点
众所周知,动物发育的最早时期在基因表达和蛋白序列进化方面都是最分散的,下一步作者统计了各物种m5C修饰丰度、基因功能区修饰位点和靶基因种类的进化方面变化程度,来观察各物种的胚胎发育过程的区别。在观察Type I和Type II m5C位点时作者有了两个新发现,第一个发现是从无脊椎动物到哺乳动物进化过程中5’UTR末端Type I位点从稀少到富集。为了解释第一个发现,作者首先测试是否可能是由于哺乳动物NSUN2蛋白的反式调控变化,并分别将人、小鼠、斑马鱼的NSUN2基因在NUSN2 KO的HeLa细胞中表达,结果显示所有其他物种NSUN2蛋白都与人NSUN2蛋白类似:
1.小鼠和斑马鱼NSUN2介导修饰的50%以上m5C位点与人NSUN2位点重叠,如下图;

2. 小鼠和斑马鱼NSUN2修饰位点序列和结构偏好性与人NSUN2类似,如下图;
3. 小鼠和斑马鱼NSUN2同样在5’UTR末端展示出m5C修饰偏好性,如下图;
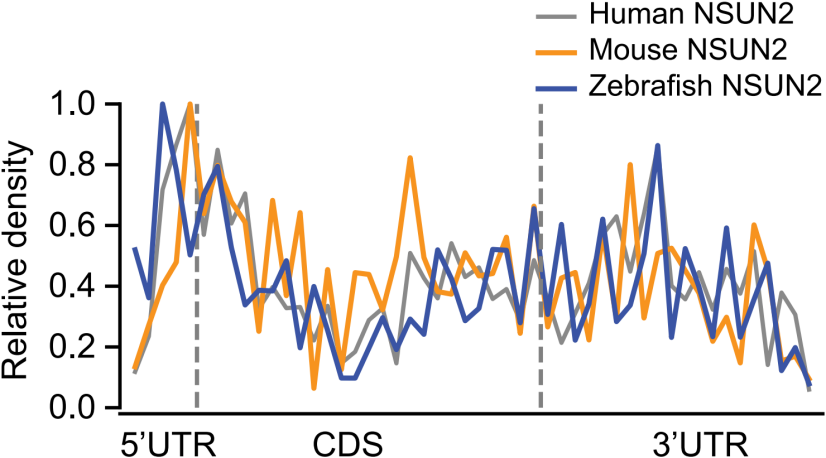
这些结果说明NSUN2反式调控变化无法解释这一发现。反式调控无法解释,只能从顺式调控入手。在物种内水平,作者发现Type I m5C位点与局部序列GC含量有关,如下图。
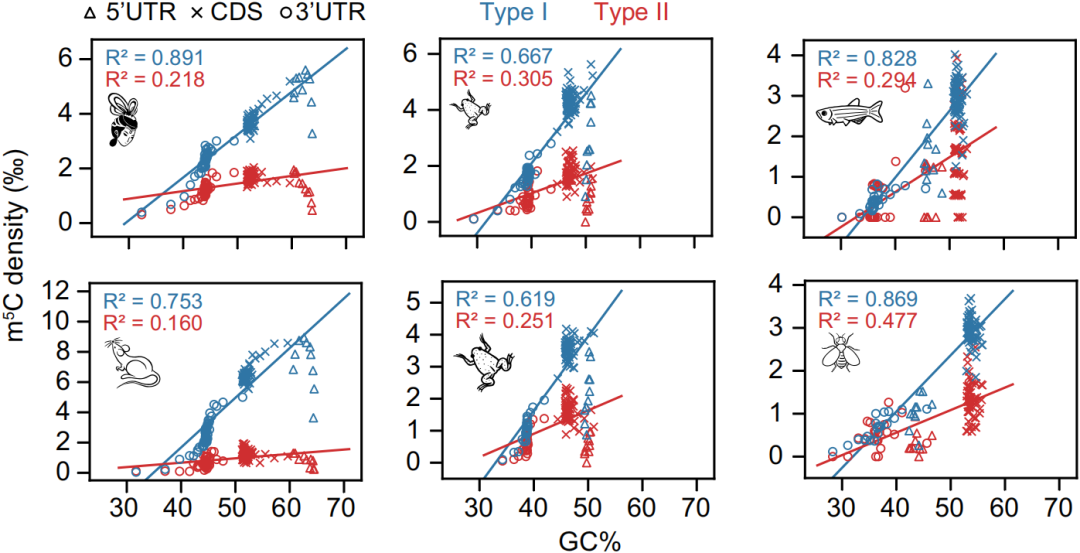
而在物种间水平,作者发现低等动物到高等动物的5’UTR末端GC含量逐渐升高,如下图,这与5’UTR末端Type I位点逐渐增加相符,并且符合结构依赖性的m5C修饰要求。

这些结果突显了基因结构进化与高等动物逐渐出现的表观转录组学之间的cross-talk。
5. 新型反式调控机制可以获得大量人类特异性Type II m5C位点
第二个发现是人类获得大量Type II m5C位点。除了Type II位点数量和比例的增加,Type II位点m5C修饰水平也高于Type I位点,如下图。这种差异在其他物种或人类受精卵mRNA中没有出现。
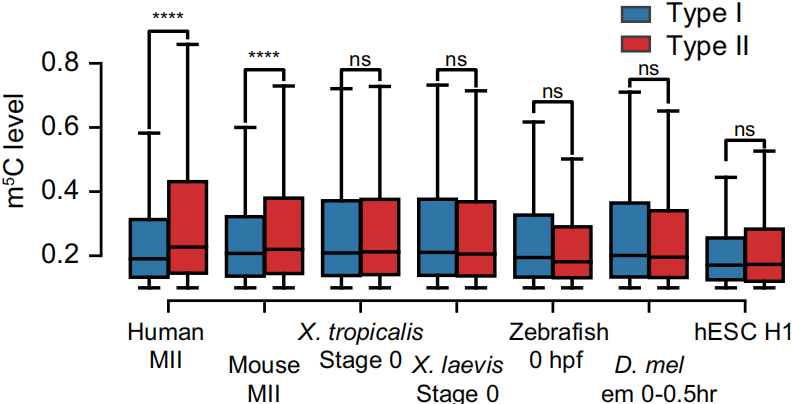
人类母体Type II位点沿转录本均匀分布,类似于其他物种和人类受精卵mRNA的情况,如下图,这说明母体Type II位点并不是人类顺式定向进化的结果。
随后发现Type II位点对mRNA二级结构的要求不高,如下图。

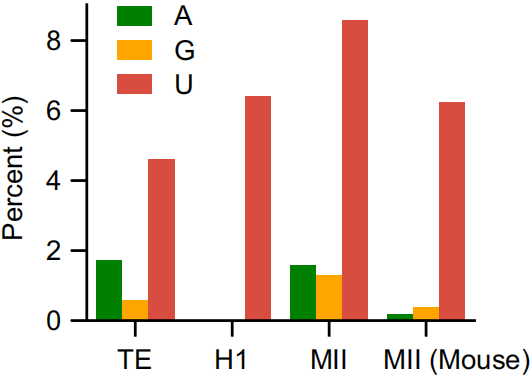
而且人类母体Type II型位点+3处的非C碱基数多于人类受精卵和其他物种m5C位点,如下图。
这些结果说明NSUN6进化到专门扩大其甲基化靶标的选择范围来调控人类母体mRNA m5C修饰。作者使用HeLa细胞模拟MI/MII卵母细胞不能显著增加Type II位点的m5C修饰水平,而且人类卵母细胞的NSUN6表达水平较高,说明NSUN6专一负责卵母细胞Type II位点m5C修饰。这些数据共同说明Type II位点在人类中的扩展可能是由于NSUN6表达和甲基化活性的反式调控的新机制。Type II位点在人类中的扩展导致约1100个基因仅由NSUN6调控甲基化,这些基因与有丝分裂细胞周期调控相关,如下图。
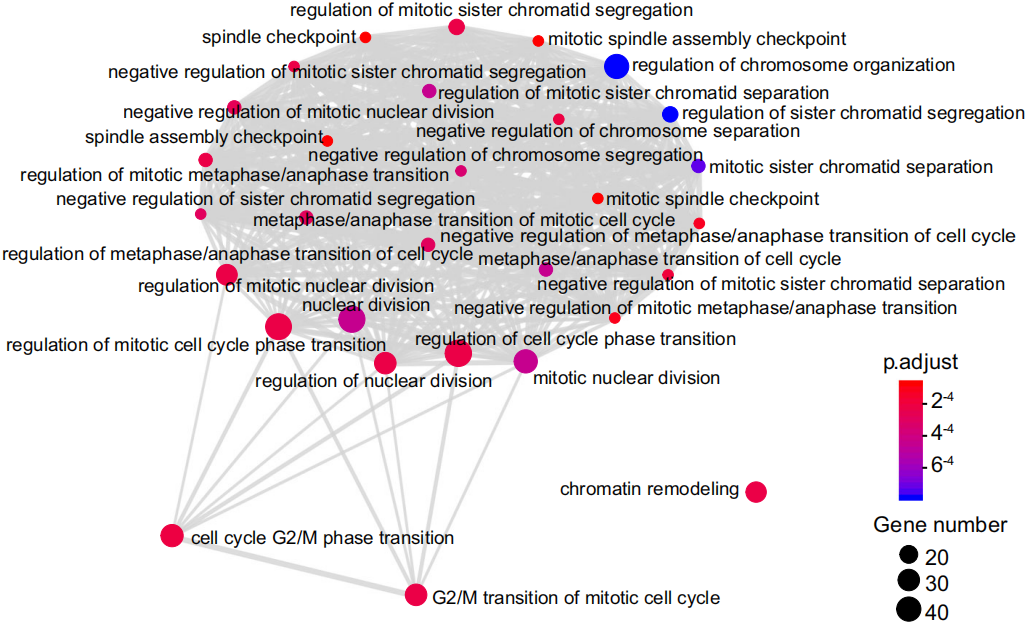
因此NSUN6介导的甲基化可能作为人类早期胚胎发育中细胞分裂调节的新机制。
6. 单个m5C位点发生快速进化
最后作者从进化角度观察m5C位点的保守性,发现随着物种偏离时间的增加,保守位点的数量和百分比在迅速下降,如下图,黑色为所有m5C位点,蓝色为Type I位点,红色为Type II位点。

鉴于单个m5C位点可以发生快速进化,作者更专注于亲缘关系较近的人和小鼠、热带爪蟾和非洲爪蟾关系对,来研究motif和结构对甲基化差异的贡献。对基序motif保守性的检测表明,在motif区域中引入的突变分别与人和小鼠、热带爪蟾和非洲爪蟾关系对中55%和37%的非保守m5C位点的甲基化丢失有关,而且保守的m5C位点都具有一个颈环二级结构,如下图。相比之下,在一个物种中被甲基化但在另一个物种中没有甲基化的位点在该物种中的茎环结构要弱很多,并且丢失甲基化,如下图。

因此,motif序列和二级结构变化都对m5C甲基化的演变做出了重大贡献。有趣的是,人类保守的Type II位点对茎环结构的要求不如小鼠严格,这与人类NSUN6进化到通过包容更多种类的二级结构和序列来扩大其甲基化靶点选择的发现相呼应。
为了定量评估motif序列和二级结构特征对m5C位点进化上的贡献,作者使用逻辑广义线性模型(GLM)分析,结果显示对于Type I位点,核心motif的+1和+4位置G碱基出现频率升高与m5C修饰水平升高有关;而+1位U碱基出现频率升高则与m5C修饰丢失有关,如下图。

对于Type II位点,除了+3位,任意核心motif位置发生突变都会导致m5C修饰的丢失,如下图。
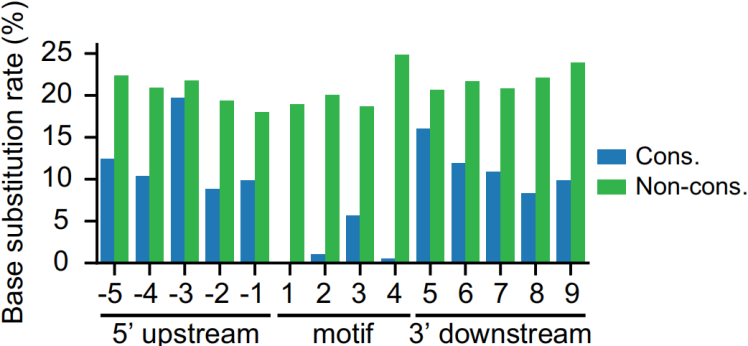
此外,尽管不同位置位点的贡献各不相同,茎环结构中配对位点的丢失和增加总体上与m5C位点的增加呈负相关,如下图。

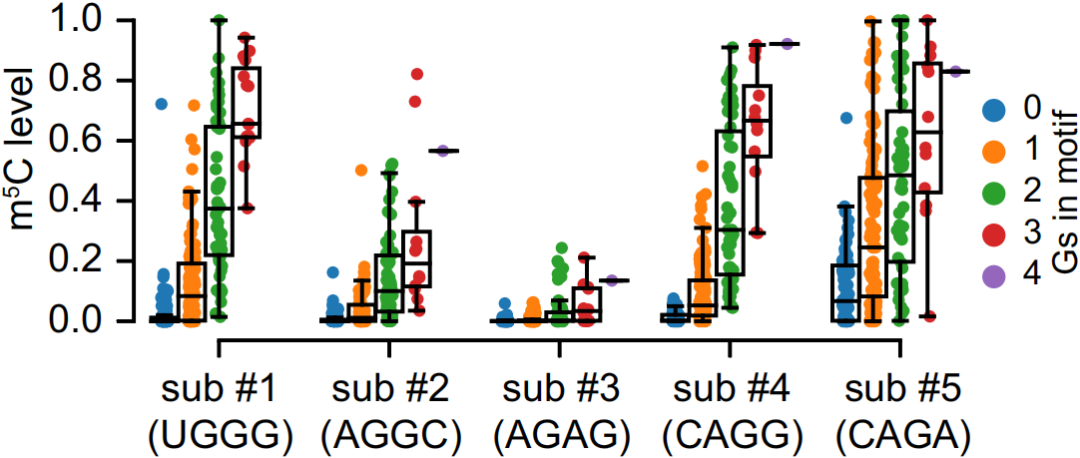
最后作者通过实验验证Type I位点3'G-rich序列中G碱基的数量与m5C修饰水平呈正相关,如下图,说明这个motif非常重要。
此外,茎环结构中,特别是m5C位点下游的五个碱基对,对于甲基化至关重要,该区域的单个突变极大地中断了甲基化修饰,如下图。
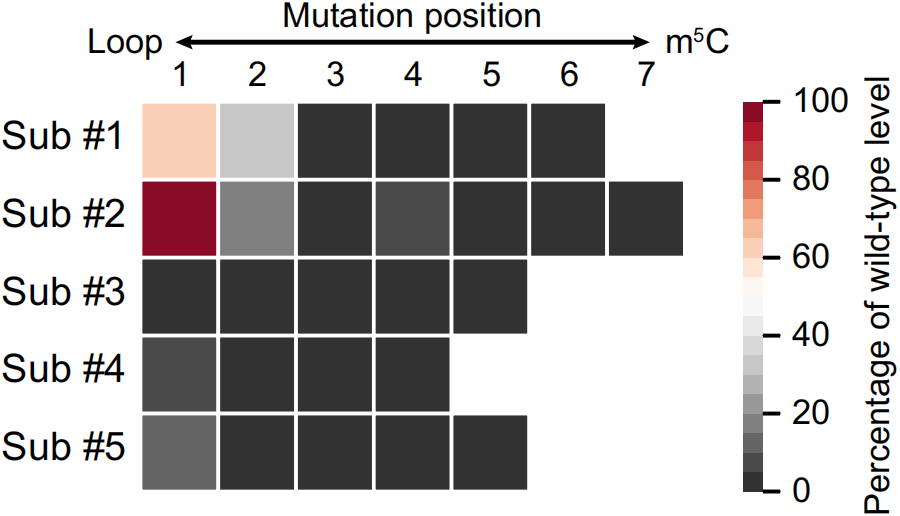
这些结果表明,m5C位点的潜在编码能力存在于周围的motif序列和茎环结构中,并且单个位点在进化过程中的增益和损失在很大程度上是可预测的。
总结
总之,这篇文章对动物发育过程中mRNA m5C的动态景观提供一个综合的视角,也对理解m5C位点的motif具有物种特异性从进化角度给出了解释。
1. 作者首先通过bsRNA-seq发现各物种母体mRNA存在大规模m5C修饰并构建m5C修饰景观;
2. 通过分析m5C motif序列并分成Type I和Type II位点,发现所有物种中Type I位点最多,而且NSUN2表达量较高;
3. 将果蝇NSUN2 KO后证明母体mRNA大规模甲基化是由NSUN2完成;
4. 从进化角度作者发现两种新机制:a. 从无脊椎动物到哺乳动物进化过程中5’UTR末端Type I位点从稀少到富集,这是由于顺式调控进化中motif序列GC含量增多所致;b. 人类获得大量Type II m5C位点,这是由于反式调控中NSUN6扩大识别序列的范围所致;
5. 进一步分析motif序列各位置碱基出现频率可以发现m5C位点在各物种间保守性较低,因此展示出物种特异性。Type I丢失一些核心位置的G碱基会导致m5C修饰水平下降;而Type II位点大部分序列突变都会导致m5C修饰水平下降,说明m5C Writers依赖motif的核心序列和二级茎环结构。

